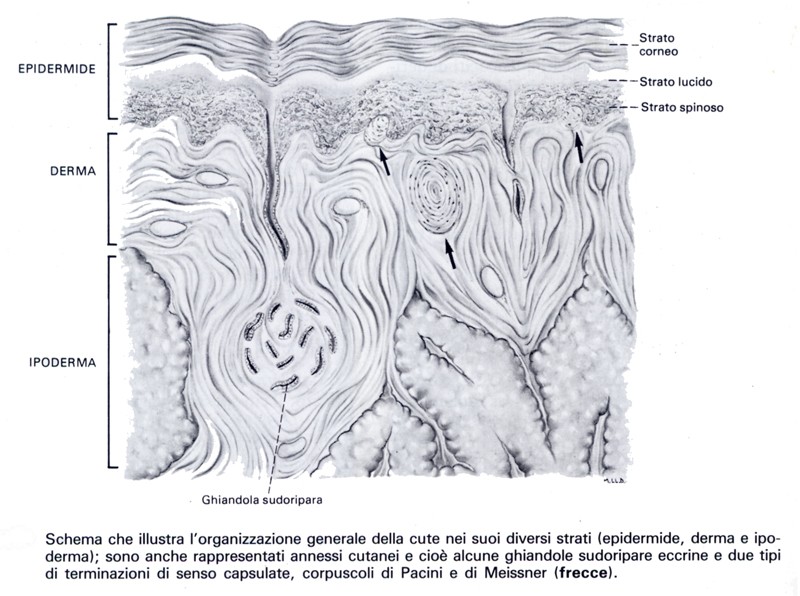
Struttura dell’organo di Corti
Struttura dell’organo di Corti L’organo di Corti rappresenta un territorio differenziato dell’epitelio di rivestimento del labirinto membranoso; esso risulta costituito dalle cellule di sostegno e dai recettori, le cellule acustiche, che sono cellule sensoriali secondarie.
Le cellule di sostegno dell’organo di Corti sono i pilastri di Corti, le cellule di Deiters, di Hensen e di Claudius.
- I pilastri di Corti sono cellule alte, rigide, impiantate sulla zona tecta della membrana basilare, dove appaiono disposte su due file (una esterna e l’altra interna). Le due strie di pilastri sono orientate in modo tale da toccarsi alla loro base d’impianto; in tal modo vengono a delimitare una galleria (di Corti). In ciascun pilastro si distinguono un piede (che poggia sulla membrana basilare), un corpo (di forma allungata) e una testa; la testa del pilastro interno è fornita di un profondo incavo che si mette in rapporto con la testa di un pilastro esterno. È da rilevare che i pilastri interni sono più numerosi di quelli esterni. Ogni pilastro è per la massima parte costituito da un materiale rigido, a struttura filamentosa, i cui filamenti sono paragonabili alle tonofibrille delle cellule epiteliali. Nel piede e nel corpo del pilastro, dal lato della galleria, si raccoglie la parte di citoplasma non differenziato, contenente il nucleo. I pilastri interni si impiantano sulla membrana basilare in vicinanza dei foramina nervina attraverso cui passano i rami terminali del nervo cocleare.
- Le cellule del Deiters sono situate esternamente ai pilastri esterni, in 3-4 file; sono cellule aventi un corpo di forma prismatica, fornito di un incavo apicale che offre sostegno a una cellula acustica esterna. Il corpo, assottigliandosi improvvisamente, continua nel collo, filiforme, assai lungo, che decorre fra le cellule acustiche esterne e termina con una piccola espansione appiattita, a forma di 8, che prende il nome di falange. Le falangi si mettono a contatto mediante i loro contorni i quali, data la particolare forma che presentano, vengono a delimitare numerosi spazi disposti regolarmente, nei quali si incastrano le parti terminali, cuticolari, delle cellule acustiche esterne. Dal contatto delle falangi delle cellule del Deiters, fra loro, con l’estremità cuticolare delle cellule acustiche esterne, con la superficie delle cellule dell’Hensen, con i pilastri esterni, e dal contatto di questi con i pilastri interni, si costituisce una superficie alla quale i contorni regolari degli elementi che si pongono a contatto conferiscono un aspetto reticolato. A tale superficie si dà il nome di membrana reticolare. Profondamente, fra i pilastri esterni e le cellule del Deiters esistono spazi intercellulari che si organizzano nello spazio di Nuel, attraversato dalle fibre nervose che dalla galleria si dirigono verso le cellule acustiche esterne. Medialmente ai pilastri interni esistono elementi di sostegno per le cellule acustiche interne, simili per la struttura alle cellule di Deiters.
- Le cellule di Hensen seguono esternamente alle cellule di Deiters. Sono elementi di sostegno, disposti in un unico strato e su più file, di altezza rapidamente decrescente procedendo dalle cellule di Deiters verso l’esterno.
- Le cellule di Claudius seguono esternamente alle cellule di Hensen; hanno l’aspetto di cellule epiteliali cubiche disposte in un unico strato e continuano lateralmente con le cellule epiteliali del solco spirale esterno, alle quali somigliano per la struttura.
Gli elementi sensoriali dell’organo di Corti sono le cellule acustiche interne e le cellule acustiche esterne, situate rispettivamente in posizione mediale e laterale rispetto alla galleria di Corti. ![]()
- Le cellule acustiche interne, circa 3500, sono disposte in un’unica fila, medialmente ai pilastri interni. Hanno aspetto cilindrico, sezione ellittica e nella loro parte apicale presentano una serie lineare di circa 20 ciglia, peli acustici, disposti a V o a U. Questi hanno la struttura delle stereociglia.
- Le cellule acustiche esterne, circa 13000, a differenza di quelle interne sono disposte su più file, da 3 a 5, passando dal giro basale a quello apicale. Hanno anch’esse forma cilindrica, ma la loro sezione è circolare e non ellittica; presentano, nella loro estremità distale, un ispessimento cuticolare dal quale si dipartono peli acustici (stereociglia) aventi disposizione analoga a quella osservata nelle cellule acustiche interne, ma assai più numerosi (60-100). A livello del polo basale di ogni cellula acustica si trovano terminazioni nervose che stabiliscono una giunzione citoneurale.
All’esame ultrastrutturale si distinguono terminazioni sinaptiche di 1° tipo, piccole e a scarso contenuto mitocondriale e vescicolare, e di 2° tipo, più grandi e ricche di vescicole. Le terminazioni a minor contenuto granulare, o piccole terminazioni nervose, hanno significato afferente; lo stimolo cioè passa dalla cellula acustica alla terminazione nervosa; le terminazioni a maggior contenuto granulare, o grandi terminazioni nervose, hanno significato efferente; ciò significa che lo stimolo passa dalla terminazione nervosa alla cellula acustica. Quest’ultima modalità di terminazione deve essere messa in rapporto con un sistema cocleare efferente che sembra svolgere funzioni inibitorie e di controllo sui recettori cocleari. La parte apicale delle cellule acustiche esterne è accolta negli spazi della membrana reticolare, delimitati dalle falangi delle cellule di Deiters.
Al di sopra della membrana reticolare e dei peli acustici è tesa la membrana tectoria, formazione cuticolare che si diparte dall’epitelio del lembo spirale; si ritiene che la membrana tectoria sia fissata, oltre che al lembo spirale, anche alla fila più esterna delle cellule di Deiters. È costituita da fibrille a direzione prevalentemente radiale, di circa 4 nm di spessore, tenute insieme da una sostanza gelatinosa ricca di mucopolisaccaridi. Il contatto tra la membrana tectoria e le ciglia delle cellule acustiche ha una grande importanza per la stimolazione dei recettori acustici. Descrivendo l’organo di Corti è importante rilevare inoltre che l’endolinfa del condotto cocleare situata al di sopra della membrana tectoria ha una composizione ionica diversa da quella dell’endolinfa contenuta negli spazi sottostanti a tale membrana: infatti, mentre la prima ha una composizione ionica con prevalenza di potassio, la seconda ha invece prevalenza del sodio, avvicinandosi quindi, sotto questo aspetto, alla composizione della perilinfa. Galleria di Corti, solco spirale interno e spazio del Nuel comunicano fra loro e sono occupati appunto da questo particolare tipo di endolinfa, cui si dà il nome di cortilinfa. La particolare composizione ionica di questo liquido viene messa in relazione con il rapporto diretto che esso ha con le fibre del nervo cocleare le quali, per potersi depolarizzare, necessitano della presenza di alte concentrazioni di ioni sodio del mezzo ambiente.


Articolo creato il 16 settembre 2012.
Ultimo aggiornamento: vedi sotto il titolo.