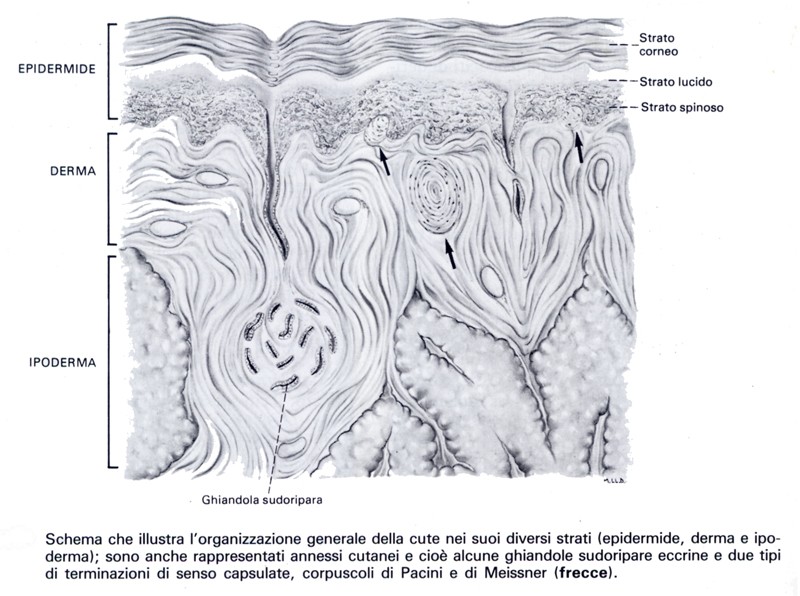
Struttura della retina
La parte ottica della retina è formata da due foglietti: uno esterno (o epitelio pigmentato) e uno interno (o retina propriamente detta). La retina propriamente detta è formata da tre strati di cellule nervose che, procedendo dall’epitelio pigmentato verso l’interno dell’occhio, si susseguono in quest’ordine: strato dei fotorecettori (cellule dei coni e dei bastoncelli), strato delle cellule bipolari e strato delle cellule multipolari.
Ciascuno di questi tre strati è ulteriormente suddiviso in altri, cosicché gli strati della parte ottica della retina risultano 10. La loro successione dall’esterno verso l’interno avviene così:
- Epitelio pigmentato.
- Strato delle cellule dei coni e dei bastoncelli.
- Membrana limitante esterna.
- Strato dei granuli esterni.
- Strato plessiforme esterno.
- Strato dei granuli interni.
- Strato plessiforme interno.
- Strato delle cellule multipolari.
- Strato delle fibre nervose.
- Membrana limitante interna.
In aggiunta ai neuroni già citati che formano con le loro diverse parti ben 7 dei 10 strati enumerati, si devono poi considerare le cellule di associazione e le cellule di sostegno che sono presenti in alcuni di questi strati.
L’epitelio pigmentato agisce non solo come schermo idoneo ad assorbire i raggi luminosi ed evitarne quindi la riflessione, ma interviene probabilmente anche nella formazione dei pigmenti visivi e negli scambi nutritizi tra lo strato coriocapillare della corioidea e i fotoreccetori. L’epitelio pigmentato si estende per tutta la parte ottica della retina, esclusa la papilla ottica; esso prosegue nel foglietto esterno delle parti ciliare e iridea della retina.
L’epitelio pigmentato è composto da un unico strato di elementi prismatici, a base esagonale, poco alti, che poggiano su una membrana basale facente parte della lamina vitrea della corioidea. Il corpo di queste cellule presenta superfici regolari laddove è a contatto con le cellule adiacenti e laddove è in rapporto con la membrana basale: regolare appare, invece, la sua estremità interna, in rapporto coi fotorecettori, per la presenza di numerose frange che si dispongono attorno al segmento esterno dei coni e dei bastoncelli. Le cellule dell’epitelio pigmentato sono particolarmente ricche in granuli di melanina, soprattutto nella loro estremità interna e nelle frange, mentre sono quasi sprovviste nella zona basale dove ha sede il nucleo.
Cellule dei coni e cellule dei bastoncelli sono le parti esterne, differenziate, delle cellule visive della retina (fotorecettori). I fotorecettori sono neuroni altamente specializzati nei quali, per effetto della luce, inizia una complessa serie di reazioni chimiche e chimico-fisiche che si traducono nella produzione di un impulso nervoso il quale viene poi trasmesso lungo particolari vie nervose fino alla corteccia telencefalica. Le cellule dei coni sono concentrate nella regione maculare e in particolare nella fovea; sono in numero di circa 5 milioni. Le cellule dei bastoncelli occupano le restanti porzioni della retina e sono in numero di circa 100 milioni.
A livello del mioide, il plasmalemma dei fotorecettori subisce alcune modificazioni consistenti in una serie di complessi giunzionali che uniscono cellule contigue. L’estremità esterna, ispessita, delle cellule di sostegno (gliociti o cellule di Muller) si mette pure in rapporto con queste giunzioni disponendosi nel loro stesso piano e allineandosi con esse: giunzioni ed estremità esterne delle cellule del Muller formano così la membrana limitante esterna che in microscopia ottica appare come una membrana anista.
L’articolo interno dei fotorecettori continua in una parte ristretta, la fibra esterna, che è esilissima nelle cellule dei bastoncelli, mentre nelle cellule dei coni appare come un breve restringimento del corpo cellulare appena accennato. Alla fibra esterna, che contiene qualche ribosoma e formazioni del reticolo endoplasmatico liscio segue una parte rigonfia delle cellule (il pericario) nella quale è contenuto il nucleo di diametro di 5-6 mm. I nuclei dei fotorecettori sono disposti in 6-76 strati, occupano uno spessore di circa 30 mm e formano lo strato dei granuli esterni.
Al di sotto del nucleo, il corpo del fotorecettore si assottiglia assumendo l’aspetto di un fine neurite amielinico contenente rari filamenti e mitocondri. L’espansione terminale di questo neurite che si mette in sinapsi con i dendriti delle cellule bipolari è diverso nelle cellule dei bastoncelli e dei coni. Nei primi il terminale simpatico, sferula, è scavato al centro da una profonda doccia nella quale penetra l’espansione dendritica della cellula bipolare. Nei secondi il rapporto fra il terminale presinaptico, pedicello, e l’elemento postsinaptico non è così stretto.
Lo strato plessiforme esterno, privo di nuclei, è formato dai neuriti fotorecettori e dai dispostivi sinaptici dei loro terminali con i dendriti delle cellule bipolari. Vi si trovano inoltre dendriti e neuriti delle cellule orizzontali.
Lo strato dei granuli interni contiene il corpo cellulare di numerosi neuroni, le cellule bipolari, il dendrite delle quali prende rapporto nello strato plessiforme esterno, con i terminali sinaptici delle cellule dei coni e dei bastoncelli il cui neurite si mette in sinapsi, nello strato plessiforme interno, con i dendriti e il pirenoforo delle cellule multipolari. Le cellule bipolari possono essere distinte in due tipi: cellule bipolari di tipo monosinaptico, il cui dendrite è in collegamento con il pedicello di un solo cono e il neurite con una cellula multipolare; cellule bipolari di tipo multisinaptico, il cui dendrite si collega con più fotorecettori e il cui neurite è in rapporto con una o più cellule multipolari. Esistono inoltre cellule bipolari invertire i cui dendriti sono rivolti verso le cellule multipolari mentre il neurite contrae sinapsi con il fotorecettore. Nella parte più superficiale dello strato dei granuli interni sono anche presenti cellule orizzontali, associative, che hanno la funzione di stabilire un collegamento fra gruppi di fotorecettori: il loro dendrite si mette in sinapsi solo con il pedicello dei coni, mentre il neurite, ramificandosi, si unisce alle terminazioni sinaptiche dei coni e dei bastoncelli. Nella parte profonda di questo strato si trovano infine le cellule amacrine, voluminosi neuroni associativi, in molti dei quali non è dimostrabile il neurite. I dendriti delle cellule amacrine contraggono rapporto con i dendriti delle cellule multipolari nel sottostante strato plessiforme interno. Lo strato dei granuli interni accoglie il nucleo delle cellule di Muller.
Lo strato plessiforme interno è l’intreccio di fibre formato dai neuriti delle cellule bipolari, che si pongono in rapporto sinaptico con i dendriti delle cellule multipolari, e dalle ramificazioni dendritiche delle cellule amacrine. Nello strato plessiforme interno possono trovarsi, sparse qua e là, in scarso numero, cellule amacrini in posizione profonda.
Lo strato delle cellule multipolari è occupato dai pirenofori di grossi neuroni multipolari (impropriamente denominati anche gangliari) disposti in un unico piano; con essi prendono pure rapporto sinaptico i neuriti discendenti delle cellule bipolari.
Lo strato delle fibre nervose è costituito dai neuriti (amielinici) delle cellule multipolari che decorrono seguendo la superficie interna, concava, del bulbo oculare fino a convergere nella papilla ottica. Qui passano a formare le fibre della parte intrabulbare del nervo ottico, riunendosi in fascetti che penetrano nei fori della lamina cribrosa della sclera.
La membrana limitante interna è formata dall’unione delle parti basali, espanse e tra di loro contigue, delle cellule del Muller. Queste sono elementi di natura gliale, molto sviluppate in altezza, dei quali, attraversando tutti gli strati della retina, giungono fino alla membrana limitante esterna. Svolgono quindi una funzione di sostegno per i vari tipi di neuroni retinici. Circa a metà della loro altezza, nello strato dei granuli interni, si trova il nucleo.

Articolo creato il 16 settembre 2012.
Ultimo aggiornamento: vedi sotto il titolo.