Formazione dell’urina
Il primo stadio della formazione dell’urina è l’ultrafiltrazione del plasma che avviene nel corpuscolo renale, attraverso i capillari glomerulari. Per ultrafiltrazione s’intende il movimento passivo di un liquido, essenzialmente privo di proteine, dal capillare glomerulare alla capsula di Bowman. L’ultrafiltrato glomerulare (o urina primaria) è prodotto nella quantità media di circa 180 litri nelle 24 ore.
Il processo di filtrazione glomerulare è reso possibile dall’elevata pressione idrostatica (50 mm/Hg) del sangue all’interno dei capillari glomerulari; a tale pressione si oppone quella esistente nello spazio capsulare (circa 10 mm/Hg) e la pressione oncotica esercitata dalle proteine plasmatiche (circa 30 mm/Hg). Ne consegue che la pressione utile di filtrazione si aggira di norma sui 10 mm/Hg. Il risultato di questi complessi meccanismi è rappresentato dall’ultrafiltrato glomerulare che nella quantità di circa 180 litri nelle 24 ore si raccoglie negli spazi capsulari e viene convogliato nei tubuli prossimali.
In conclusione, l’ultrafiltrazione è regolata dalle leggi del trasporto passivo attraverso le membrane, dipende dalle caratteristiche fisiche della membrana filtrante e dalla differenza della pressione idrostatica tra le due superfici della membrana stessa, e si svolge a spese dell’energia fornita dal cuore. La lamina basale, la più impermeabile delle formazioni che compongono questa membrana filtrante, si comporta come se fosse fornita di canali lunghi 0,25-0,35 mm, corrispondente allo spessore della lamina stessa, e di un diametro inferiore a 7 nm, che è la dimensione della maggior parte delle proteine del sangue. A questo riguardo va tenuto presente, soprattutto per le proteine, che ha importanza non soltanto il peso, ai fini del loro passaggio, ma anche la loro configurazione (globulare o fibrosa) e la carica, che è in relazione con il punto isoelettrico della proteina stessa.
L’endotelio dei capillari glomerulari è di tipo fenestrato (contiene pori del diametro di circa 700 Å; 1Å = 10-10 m) ed è liberamente permeabile all’acqua, ai piccoli soluti come il sodio, all’urea e al glucosio, e persino a piccole molecole proteiche, ma è impermeabile alle cellule e a sostanze di elevato peso molecolare, quali le proteine plasmatiche; solamente l’albumina compare in piccola quantità nel filtrato glomerulare, ma viene prontamente riassorbita nel tratto successivo del nefrone, il tubulo prossimale. La matrice basale, che è una matrice porosa di proteine extracellulari (tra cui la laminina, la fibronectina, il collageno di tipo IV) costituisce un’importante barriera per la filtrazione delle proteine plasmatiche. I podociti possiedono processi digitiformi tra loro interdigitati e separati da fessure dette fessure di filtrazione. Queste strutture rallentano la filtrazione di alcune proteine e macromolecole che riescono a passare attraverso l’endotelio e la membrana basale. Poiché le cellule endoteliali, la membrana basale e le fessure di filtrazione contengono glicoproteine cariche negativamente, la parete del capillare glomerulare filtra le sostanze in base sia alle loro dimensioni che alla loro carica elettrica. Per molecole che abbiano un raggio molecolare compreso tra 20 e 42 Å, quelle cationiche sono filtrate più facilmente di quelle anioniche. Inoltre, i podociti sono in grado di effettuare l’endocitosi che permette al materiale di entrare nella cellula senza attraversare la membrana.
L’endotelio fenestrato dei capillari, la lamina basale e i podociti che formano il foglietto viscerale della capsula glomerulare costituiscono nell’insieme la cosiddetta barriera di ultrafiltrazione (o barriera sangue/urina), che nel soggetto adulto ha un’estensione complessiva di circa 0,8 m2. In questa barriera, la lamina basale rappresenta lo strato più spesso e privo di discontinuità.
L’ultrafiltrato glomerulare che si raccoglie nello spazio capsulare presenta lo stesso contenuto di elettroliti del plasma e ne ha le medesime caratteristiche chimicofisiche; si differenzia dal plasma soltanto per la virtuale assenza di proteine.
L’ultrafiltrato giunge quindi nel tubulo renale che ha la funzione di modificarlo, trasformandolo nell’urina definitiva, in seguito alle peculiari proprietà assorbenti e secernenti delle cellule epiteliali che lo delimitano. In tal modo, l’ultrafiltrato glomerulare, durante il transito nel tubulo, risulta concentrato per il riassorbimento di oltre il 99% dell’acqua e modificato nella sua composizione chimica e fisicochimica. L’ultrafiltrato glomerulare risulta così ridotto a circa 1,5 litri, che corrisponde al volume dell’urina emessa da un soggetto sano a dieta ordinaria in un giorno.
Il tubulo renale è costituito dal tubulo prossimale, dall’ansa di Henle e dal tubulo distale. Nel prendere in considerazione nella sua complessità le modificazioni che subisce l’ultrafiltrato glomerulare fino alla formazione dell’urina, si considera parte integrante del tubulo renale, e quindi del nefrone, anche il tubulo collettore. Quest’ultimo, infatti, raccogliendo più tubuli renali non viene, anche per ragioni embriologiche, considerato come parte del nefrone: da un punto di vista funzionale non si possono, tuttavia, trascurare le sue importanti attività nella uropoiesi.
La funzione principale del tubulo prossimale consiste nel riassorbire circa l’80% in volume dell’ultrafiltrato glomerulare. Oltre all’acqua, all’urea e a numerosi elettroliti, vengono riassorbite selettivamente altre sostanze (glucosio, fruttosio, galattosio, aminoacidi, albumina, ecc.) che avevano attraversato la barriera di filtrazione entrando nella composizione dell’ultrafiltrato glomerulare. Il riassorbimento può essere passivo (per diffusione) o attivo (per trasporto). In particolare, l’assorbimento degli ioni sodio avviene mediante un trasporto attivo da parte delle cellule tubulari con notevole dispendio energetico, mentre quello degli ioni cloro e dell’acqua avviene passivamente per ristabilire l’equilibrio delle cariche elettriche e per mantenere isosmotico l’ambiente intra- ed extratubulare (riassorbimento obbligato). Inoltre, il riassorbimento di aminoacidi e di esosi sembra avere luogo a livello dei microvilli apicali con un meccanismo attivo mediato dalla presenza di molecole trasportatrici (carriers). Attivamente viene riassorbita quella minima quantità di proteine a più basso peso molecolare (sieroalbumina, emoglobina) che aveva attraversato la barriera di ultrafiltrazione. Le proteine vengono assunte dalle cellule con il meccanismo dell’endocitosi, testimoniato dalla presenza di vacuoli citoplasmatici. Le cellule del tubulo prossimale, infine, secernono nella preurina numerose sostanze endogene (ioni potassio, ioni idrogeno, creatinina) ed esogene (per esempio, la penicillina e altri antibiotici). Alla fine del tubulo prossimale l’ultrafiltrato glomerulare è ridotto a circa 1/5 del suo volume iniziale e prende il nome di liquido tubulare.
Nell’ansa di Henle il liquido tubulare viene ulteriormente modificato. Nel braccio discendente subisce un processo di concentrazione per il riassorbimento di acqua e la secrezione di ioni sodio e cloro e di urea. Al contrario, nel braccio ascendente, impermeabile all’acqua, vengono riassorbiti ioni sodio e cloro (pompa del sodio); ne consegue una diminuzione della concentrazione del liquido tubulare senza apprezzabili modificazioni del suo volume per il mancato riassorbimento di acqua. I processi che si svolgono nell’ansa del nefrone hanno dato origine all’ipotesi della concentrazione osmotica controcorrente che deriva dalla particolare disposizione delle anse e dei vasi retti nella zona midollare. Ammettendo che in un tubo ad ansa circoli una soluzione salina e che le pareti contigue delle anse abbiano la capacità di trasferire i sali disciolti, la pompa del sodio permetterà la fuoriuscita di questo ione lungo tutto il braccio ascendente dell’ansa rendendo così iperosmotico il liquido interstiziale, mentre non consentirà il passaggio dell’acqua. Conseguentemente, con meccanismo di diffusione e quindi passivo, uscirà acqua soltanto dal braccio discendente dell’ansa stessa, mentre vi penetreranno con lo stesso meccanismo di diffusione ioni sodio e cloro e parzialmente anche l’urea; pertanto il liquido di questo tratto diventerà sempre più concentrato rispetto al liquido contenuto nel braccio ascendente.
Essendo però quest’ultimo impermeabile all’acqua, il liquido in esso contenuto sarà sempre più diluito fino a divenire ipotonico. In seguito l’effetto dell’ormone antidiuretico (o vasopressina ADH), intervenendo sull’epitelio della parte convoluta dei tubuli distali e soprattutto dei tubuli collettori, consentirà il riassorbimento dell’acqua e quindi la concentrazione definitiva dell’urina. Infatti, l’urina eliminata presenta la concentrazione osmotica del liquido interstiziale che si trova all’apice delle papille renali, corrispondente a quattro volte la concentrazione osmotica del plasma e quindi dell’ultrafiltrato glomerulare. In conclusione, lo scambio osmotico controcorrente è un processo passivo che non richiede dispendio energetico oltre quello necessario alla circolazione del liquido tubulare nell’ansa eseguito a spese del lavoro del cuore. Il gradiente osmotico a livello delle piramidi midollari non permarrebbe a lungo se lo ione sodio, gli altri ioni e l’urea, che rendono iperosmotico il liquido interstiziale, venissero asportati dalla circolazione. Al loro mantenimento provvedono i vasi retti che funzionano da moltiplicatori osmotici controcorrente.
Nel segmento iniziale rettilineo del tubulo distale ha luogo un riassorbimento attivo di ioni sodio (tramite la pompa del sodio) i quali, passando nel liquido interstiziale del connettivo peritubulare, vi determinano un aumento della pressione osmotica, creando le basi per un ulteriore riassorbimento di acqua da parte dell’epitelio tubulare.
Nel segmento convoluto del tubulo distale, sotto l’influenza dell’ormone antidiuretico, viene riassorbita acqua (riassorbimento facoltativo) e l’urina tende a concentrarsi. Sempre in questa porzione del tubulo distale proseguono, per l’azione dell’aldosterone, il riassorbimento attivo di sodio e la contemporanea secrezione di potassio e di idrogenioni. Si ritiene, inoltre, che l’epitelio del tubulo distale produca callicreina, un enzima che passando nel sangue determina indirettamente la formazione di bradichinina.
Articolo creato il 14 agosto 2011.
Ultimo aggiornamento: vedi sotto il titolo.

Potrebbe anche piacerti:
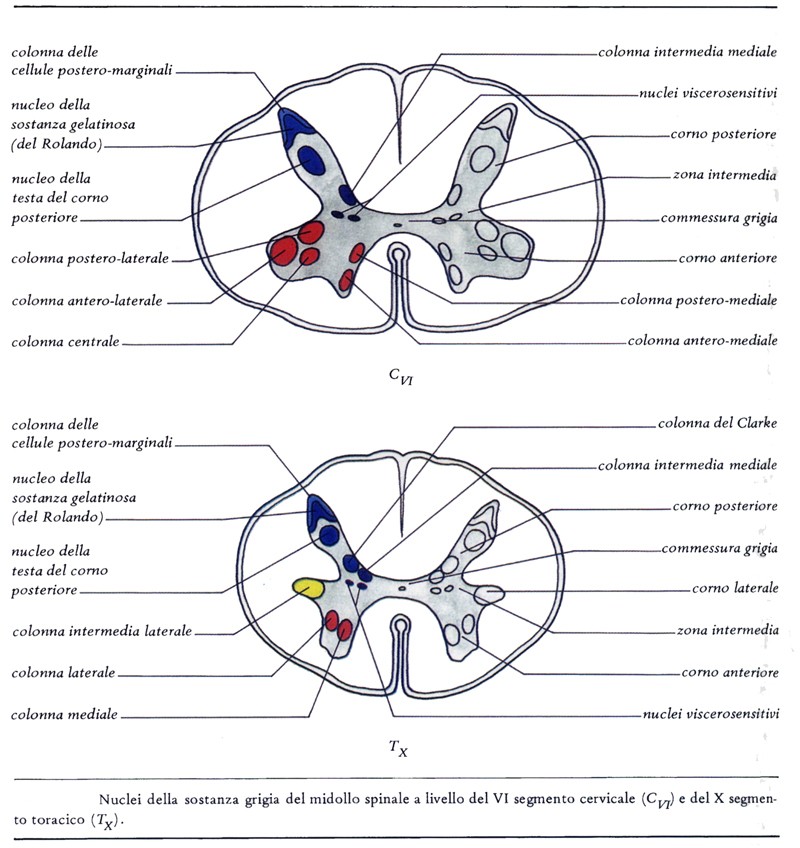
Nuclei del corno anteriore del midollo spinale

Examen oficial MIR 2008